Akrozomální reakce
Akrozomální reakce (také akrozomová reakce nebo akrosomální/akrosomová exocytosa) je exocytóza akrozomu (specifické organely spermií), která usnadní spermii proniknout glykoproteinovým obalem vajíčka – zonou pellucidou a umožní spojit se s vajíčkem (fúze gamet). U savců mohou akrozomovou reakci mohou podstoupit jen kapacitované spermie (spermie, které pobývaly určitou dobu v samičím reprodukčním traktu, dokud plně nedozrály)
-
 Akrozom
Akrozom -
 akrozomální reakce
akrozomální reakce -
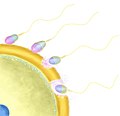 akrozomální reakce
akrozomální reakce



Předpokládaný průběh akrozomální reakce
editovat- Kontakt se zonou pellucidou vyvolá výrazné zvýšení koncentrace Ca 2+ v cytoplazmě spermie (vyšší než v průběhu kapacitace).
- Během akrozomální reakce dochází k depolymeraci F-aktinu na G-aktin, což zpřístupní cytoplasmatickou membránu spermie vnější akrosomální membráně a umožní fúzi.[1],[2]
- Dochází k fúzi cytoplasmatické membrány a vnější akrozomální membrány, objevují se tzv. fúzní póry a obsah akrozomu se dostává ven. Je buď volný anebo obalený membránou vzniklou fúzí
- Odhalí se vnitřní akrozomální membrána, která obsahuje proteiny vázající zonu pellucidu a umožní spermii udržet se na místě. Dále nese proteiny, které zprostředkují fúzi spermie s cytoplazmatickou membránou vajíčka
- Fúze membrán při akrozomální reakci
-

-
 obtočení T- a V-snare kolem sebe a přitažení membrán
obtočení T- a V-snare kolem sebe a přitažení membrán -
 vznik fúzního póru
vznik fúzního póru



Obálky vajíčka
editovatU většiny druhů savců má vajíčko dvě obálky – shluk granulózních buněk corona radiata a glykoproteinový obal – zonu pellucidu. Právě kontakt se zonou pellucidou je přirozeným spouštěčem akrozomální reakce. Samotná zona pellucida se skládá převážně ze tří glykoproteinů – ZP1, ZP2 a ZP3, přičemž se předpokládá, že právě ZP3 je hlavním stimulátorem. Nicméně sám o sobě (bez ZP1 a ZP2) akrozomální reakci chopen vyvolat není.
Regulace akrosomální reakce
editovatAkrozomální reakce je příkladem děje kontrolovaným poměrně složitými signalizačními kaskádami, jejichž vzájemné propojení a interakce dodnes nejsou plně objasněny. Akrozomální reakce je kontrolována pomocí enzymů PKA,PKC a PLC a výrazně zvýšenou koncentraci Ca2+ a depolymerací aktinu. Celý regulační proces navazuje a úzce souvisí s kapacitací
Kapacitace
editovatprostředí pohlavního traktu samice způsobí:
- odstranění dekapacitačních faktorů (faktorů zamezujících kapacitaci) – např. cholesterol, glykogelin S,…
- nárůst koncentrace HCO3− a Ca2+, postupný nárůst koncentrace ROS
- aktivace HCO3– dependentní adenyly cyklázy
- nárůst konc. cAMP
- aktivace PKA (pomocí cAMP) ( a PKC (pomocí Ca2+)
PKA modifikuje a aktivuje:
- aktivace PLD1
- PLD1 vytváří PA (kyselina fosfatidová), která nepřímo stimuluje polymeraci aktinu (interakce s proteiny reagujícími s aktinem)
- přesun PLC do cytoplazmatické membrány
- protein tyrosinová fosforylace proteinů v bičíku
Akrozomová reakce
editovatinterakce se zonou pellucidou: ještě se zvýší aktivita PKA a PKC a dojde k aktivaci PLC
- dochází k výraznému nárůstu koncentrace Ca2+ (PKC aktivuje Ca2+ iontové kanály v plazmatické membráně a PKA s PLC (pomocí PI3) zase ve vnější akrozomální membráně)
- vysoká koncentrace Ca2+ vede k depolymeraci aktinu
- vysoká koncentrace Ca2+ vede k aktivaci SNARE proteinů (pomocí proteinu Rab3) a jejich interakci.
(Navázání T a V-Snare na sebe, způsobí změnu jejich konformace, prudce se kolem sebe obtočí a přitáhnou membrány k sobě)
- přiblížení membrán pomocí SNARE proteinů vede k fúzi vnější akrozomální membrány s plazmatickou membránou
| zkratka | český název | anglický název | reakce |
| ROS | reaktivní kyslíkové readikály | reactive oxygen species | - |
| cAMP | cyklický adenosylnmonofosfát | cyclic adenosylmonophosphate | aktivace různých enzymů |
| PKA | proteinkináza A | proteinkinase A, cAMP dependentní proteinkináza | fosforylace proteinů na Ser/Thr |
| PKC | proteinkináza C | proteinkinase C, Ca2+ dependentní (a DAG dependentní) protein kináza | fosforylace proteinů na Ser/Thr |
| PLD1 | fosfolipáza D1 | phospholipase D1 | hydrolýza fosfatidylcholinu na kyselinu fosfatidovou (PA) a cholin |
| PLC | fosfolipáza C | phospholipase C | fosfatidylinositolbisfosfátu (PIP2) na inositoltrifosfát (PI3) a diacylglycerol (DAG) |
Fúze gamet
editovatOdhalená vnitřní akrozomální membrána na sobě nese specifické proteiny (proteinové receptory – z proteinové rodiny ADAM), které jsou schopné interagovat s jinými proteiny na plazmatické membráně vajíčka (integriny, CD9). Touto interakcí dojde k přiblížení a nakonec i spojení těchto membrán – tedy k fúzi gamet.
Enzymy akrozomu
editovatObsah akrozomu tvoří různé lytické enzymy:
- proteázy: akrosin (proteáza trypsinového typu exprimovaná pouze ve spermatogenních buňkách) a jeho prekurzor preakrosin, kalpain II, pepsidáza podobná kolagenáze, pepsidáza podobna katepsinu D
- glykosidázy: β-galaktosidáza, hyaluronidáza, neuraminidáza, β-N-acetylglukosaminidáza
- lipázy: fosfolipáza A2, fosfolipázy C
- další – arylsulfatáza, …
Předčasná akrozomální reakce
editovatnebo také nespecifická akrozomální reakce. V průběhu kapacitace (či po ní) některé spermie podstoupí zonou pellucidou nevyvolanou akrozomální reakci a uvolní tak obsah akrozomu. Předčasně akrozomově zreagované spermie (jinak označované za spermie s porušených/ chybějícím akrozomem) mají výrazně sníženou šanci oplodnit vajíčko.
Akrozomální reakce in vitro
editovatPři výzkumu oplození a plodnosti (neboli fertilizace a fertility) je často zapotřebí vyvolat akrozomovou reakci . Proteiny zony pellucidy (ZPp) nejsou standardně komerčně dostupné (obzvláště lidské) . Důvodem je nefunkčnost transgenních ZPp. Využívá se tedy jiných látek, které jsou schopné akrozomovou reakci vyvolat uměle. Nejčastější je použití ionoforu Ca2+ (např. A23187 nebo ionomycin), který zprůchodní membránu pro Ca2+ a způsobí tak výrazné zvýšení cytoplazmatické koncentrace tohoto iontu. Dále se používá progesteron, který také dokáže zvýšit koncentraci cytoplazmatického Ca2+.[6] Vysoká koncentrace Ca2+ u kapacitované spermie pak vede k akrozomové reakci. Úspěšnost akrozomální reakce lze dokázat běžnými histologickými barveními (např. May-Grünwaldovo barvení) nebo lektinovými studiemi (např. pomocí WGA, ConA) anebo protilátkami proti specifickým akrozomálním proteinům (např. akrosin) (akrozomálně zreagované spermie už tyto proteiny nemají, popř. pouze vázané na vnitřní akrozomální membránu)
Příklad vyvolání akrozomové reakce in vitro
editovatK suspenzi kapacitovaných spermií v kapacitačním médiu se přidá A 23187 (konc. 10µM) a spermie se nechají inkubovat při T = 37 °C, 5% CO2 atmosféře a 100% vlhkosti vzduchu po dobu 30min.[7]
Související články
editovatOdkazy
editovatReference
editovat- ↑ Brener E, Rubinstein S, Cohen G, Shternall K, Rivlin J, Breitbart H, Remodeling of the actin cytoskeleton during mammalian sperm capacitation and acrosome reaction. Biol Reprod. 2003 Mar;68(3):837-45
- ↑ Breitbart H, Cohen G, Rubinstein S. Role of actin cytoskeleton in mammalian sperm capacitation and the acrosome reaction. Reproduction. 2005 Mar;129(3):263-8
- ↑ Breitbart H. „Signaling pathways in sperm capacitation and acrosome reaction.“ Cell Mol Biol (Noisy-le-grand). 2003 May;49(3):321-7.
- ↑ a b Ramalho-Santos J, Schatten G, Moreno RD.“ Control of membrane fusion during spermiogenesis and the acrosome reaction.“ Biol Reprod. 2002 Oct;67(4):1043-51.
- ↑ Breitbart H, Cohen G, Rubinstein S. Role of actin cytoskeleton in mammalian sperm capacitation and the acrosome reaction. Reproduction. 2005 Mar;129(3):263-8.
- ↑ Brucker C, Lipford GB. „The human sperm acrosome reaction: physiology and regulatory mechanisms. An update.“ Hum Reprod Update. 1995 Jan;1(1):51-62.
- ↑ Mendoza C, Carreras A, Moos J, Tesarik J., „Distinction between true acrosome reaction and degenerative acrosome loss by a one-step staining method using Pisum sativum agglutinin.“ J Reprod Fertil. 1992 Aug;95(3):755-63.
Literatura
editovat- Neil JD, „Knobil and Neill's Physiology of Reproduction“, Academic Press; 3 edition (12 Dec 2005), ISBN 0125154003